Видообразуване (специация): как се формират нови видове в еволюцията
Видообразуване (специация): научно обяснение как се формират нови видове — от географска изолация до хибридизация и ДНК анализи, примери и механизми на еволюцията.
Видообразуването е свързано с начина на формиране на видовете. То е основна част от еволюционната биология и обяснява как от една популация постепенно се появяват две или повече популации, които вече не обменят гени помежду си и имат различни наследствени характеристики.
Дарвин е смятал, че повечето видове произлизат директно от вече съществуващи видове. Това се нарича анагенеза — промяна на един вид във времето (филетична еволюция). През по-голямата част от XX век учените смятаха, че повечето видове се появяват чрез разделяне на по-ранни видове — процес, известен като кладогенеза. Общото мнение беше, че видовото разделяне обикновено се случва чрез развитие на различни изолиращи механизми, които пречат на размяната на гени между популациите.
Галерия с изображения
10 Изображения









Основни причини и механизми
Без съмнение един от най-важните фактори е физическото разделяне на популации, които някога са живели заедно. Това води до прекъсване на генния поток и позволява на различните популации да натрупват различия чрез мутация, естествен подбор и генетичен дрейф.
Работата през последните десетилетия показа, че има и други причини за видообразуване. Анализът на ДНК последователността на живите организми показа, че често има хибридизация между сродни видове — т.е. при кръстосване се пренасят гени от единия вид в друг. Това показва, че репродуктивната изолация не винаги е абсолютен критерий за вид и че не винаги е необходима пълна алопатрия (географска изолация) за възникване на нови видове.
Видове изолация
Различните изолационни механизми, които водят до намален или прекратен генен поток, се групират по принцип:
- Презиготни (пред-оплождащи) механизми: различно време на размножаване, различно поведение/полови предпочитания, механична несъвместимост на половите органи, невъзможност за привличане на партньор (например специфични сигнали или миризми).
- Постзиготни (след-оплождащи) механизми: намалена жизнеспособност на хибридите, стерилност (например муле), или намалена плодовитост в последващи поколения.
Основни форми на видообразуване
- Алопатрично видообразуване — когато популации се разделят географски (реки, планини, острови) и се еволюират независимо. Класически пример са островните видове, например Дарвиновите врабчета.
- Периаптрично и паропатрично видообразуване — при периаптрично една малка изолирана популация (фондаторски ефект) се отклонява от главната популация; при паропатрично популациите са съседни и изолацията е частична, често свързана с градиенти в средата.
- Симпатрично видообразуване — възниква без ясно географско разделяне; може да бъде резултат от силна екологична дивергенция, сексуален подбор или генетични промени като полиплоидия при растенията.
- Полиплоидна спeцијация — особено честа при растенията: внезапно увеличение на броя на хромозомите (напр. алополиплоидия или автополиплоидия) води до репродуктивна изолация от родителската популация. Примери: някои културни видове пшеница и родове като Tragopogon.
- Хибридна спeцијация — при която хибридизацията между два вида създава нов таксономично различим и репродуктивно изолиран потомък. Това може да бъде и хомоплоидно (без промяна в хромозомния брой) или чрез полиплоидия.
Примери от природата
- Дарвиновите врабчета (Галапагос): класически пример за алопатрична и екологична дивергенция, при която различни острови и хранителни ниши довеждат до различни форми на човки и поведение.
- Африканските цихлиди: бурна радиация в езерата (напр. езеро Виктория) — множество видове, които са се формирали бързо чрез екологична адаптация и сексуален подбор.
- Саламандрите от родa Ensatina: пример за "пръстеновидни видове" (ring species), където генетичната и морфологична промяна на населване по континентален пръстен води до популации, които постепенно се различават.
- Растения: много видове възникват чрез полиплоидия и хибридизация — примери включват видове в родовете Spartina, Tragopogon и други.
- Хибридна спeциация при слънчогледи (род Helianthus): известни случаи, при които хибриди са дали началото на нови адаптирани видове.
Роля на генетиката и молекулярните данни
Молекулярните изследвания (сравняване на ДНК последователности) позволиха да се видят сложните връзки между популации и видове. Те показаха, че:
- генният поток (миграция на гени) понякога продължава дълго, въпреки началната дивергенция;
- периоди на интензивна хибридизация могат да внесат адаптивни гени от един вид в друг (introgression);
- има "континуум" от популации — от слабо различими локални форми до напълно отделни видове.
Темпо и модел на видообразуване
Учeните дискутират дали видовете възникват постепенно (градуализъм) или в бързи периоди, последвани от относителна стабилност (punctuated equilibrium). Днес повечето изследователи признават, че и двете схеми са възможни в зависимост от причините за дивергенция (напр. бърза екологична промяна, колонизация на нова среда, полиплоидия).
Какво означава това за понятието "вид"?
Съвременните данни показват, че границите между видовете могат да бъдат размити. Различни дефиниции на "вид" се използват в зависимост от контекста: биологична дефиниция (репродуктивна изолация), морфологична, филогенетична и др. В практиката често се комбинират генетични, морфологични и екологични данни, за да се реши дали дадени популации представляват различни видове.
Накратко, видообразуването е многоаспектен процес, в който участват география, екология, генетика, поведение и случайни фактори. Физическото разделяне е силно влияние, но не единствено — хибридизацията, полиплоидията и екологичната дивергенция също могат да доведат до появата на нови видове.
Изолиращи механизми
Изолиращите механизми са неща, които възпрепятстват успешното размножаване между групите от даден вид. Установява се репродуктивна изолация на популациите. Това е особено важно за концепцията за биологичните видове, тъй като видовете се определят чрез репродуктивна изолация.
Изолиращите механизми могат да бъдат разделени на две групи - преди и след оплождането.
Преди оплождане
Фактори, които пречат на индивидите да се чифтосват.
- Географска изолация: Видовете се срещат в различни райони и често са разделени от бариери.
- Времева изолация: Индивидите не се чифтосват, защото са активни по различно време. Това може да бъде различно време на деня или различни сезони. Възможно е периодите на чифтосване на видовете да не съвпадат. Индивидите не се срещат един с друг по време на периодите на чифтосване или изобщо не се срещат.
- Екологична изолация: Индивидите се чифтосват само в предпочитаното от тях местообитание. Те не се срещат с индивиди от други видове с различни екологични предпочитания.
- Поведенческа изолация: Индивидите от различни видове могат да се срещнат, но единият не разпознава никакви сексуални сигнали, които могат да бъдат дадени. В повечето случаи индивидът избира представител на своя вид.
- Механична изолация: Може да се направи опит за копулация, но не се осъществява трансфер на сперматозоиди. Индивидите може да са несъвместими поради размера или морфологията си.
- Геймическа несъвместимост: Преносът на сперматозоиди се осъществява, но яйцеклетката не се опложда.
След оплождане
Фактори, които пречат на успешното чифтосване, като генетична несъвместимост, хибридна нежизнеспособност или стерилитет.
- Зиготна смъртност: Яйцеклетката е оплодена, но зиготата не се развива.
- Хибридна нежизнеспособност: Хибридният ембрион се формира, но не е жизнеспособен.
- Хибриден стерилитет: Хибридът е жизнеспособен, но полученото възрастно е стерилно.
- Хибридна разбивка: Първото поколение хибриди (F1) е жизнеспособно и плодовито, но следващите поколения хибриди (F2 и обратни кръстоски) са нежизнеспособни или стерилни.

Географска изолация
Смята се, че това е най-честата причина за видообразуване. Първият човек, който се сеща за нея, е Мориц Вагнер, немски изследовател и естествоизпитател.
В началото на кариерата си Вагнер е географ и публикува редица географски книги за Северна Африка, Близкия изток и Тропическа Америка. Освен това той е запален естествоизпитател и колекционер и именно с тази си дейност е най-известен сред биолозите. Ернст Майр, еволюционистът и историкът на биологията, разказва за значението на Вагнер.p562–565
По време на тригодишния си престой в Алжир Вагнер (наред с други дейности) изучава нелетящите бръмбари Pimelia и Melasoma. Всеки род се разделя на няколко вида, всеки от които е ограничен в участък от северното крайбрежие между реките, които се спускат от Атласките планини към Средиземно море. Щом се пресече река, се появява различен, но близкородствен вид.
"... един зараждащ се вид ще [възникне] само когато няколко индивида пресекат границите на своя ареал... формирането на нова раса никога няма да успее... без продължително отделяне на колонистите от останалите представители на техния вид".
Това е ранно описание на процеса на един вид географско видообразуване. През 1942 г. то е въведено отново от Майр и значението на географското видообразуване се превръща в една от основните идеи на еволюционния синтез.
Друг термин за географско видообразуване е алопатрично видообразуване. Алопатричен означава "различна земя".
Свободни острови
Вулканичните острови са образувани без живот и целият живот трябва да пристигне, носен от вятъра или водата. От Хавайските острови и от Галапагоските острови знаем, че всички форми на живот се променят, когато достигнат островите от континента.
На площ от около 6500 кв. мили (17 000 км ) на 2Хавайските острови се намира най-разнообразната колекция от дрозофилни мухи в света, които живеят от дъждовните гори до планинските ливади. Известни са около 800 вида хавайски дрозофилиди.
Проучванията показват ясен "поток" от видове от по-старите към по-новите острови. Има и случаи на колонизация обратно на по-стари острови и на прескачане на острови, но те са много по-редки.
По метода на радиоактивното датиране с калий/аргон настоящите острови датират от преди 0,4 милиона години (миа) (Мауна Кеа) до 10 миа (Некер). Най-старият член на Хавайския архипелаг, който все още е над морето, е атолът Куре, който може да се датира от 30 миа.
Самият архипелаг, образуван от Тихоокеанската плоча, която се движи над гореща точка, съществува от много по-дълго време, поне от креда. Хавайските острови плюс бившите острови, които сега са под водата, съставляват веригата Хавайско-императорски подводни планини, а много от подводните планини са гайоти.
Всички местни видове дрозофилиди на Хаваите очевидно произхождат от един-единствен прародителски вид, който е колонизирал островите преди около 20 милиона години. Последвалата адаптивна радиация е била стимулирана от липсата на конкуренция и голямото разнообразие от свободни ниши. Въпреки че е възможно една бременна женска да колонизира остров, по-вероятно е това да е била група от един и същи вид.
На Хавайския архипелаг има и други животни и растения, които са претърпели подобна, макар и не толкова впечатляваща, адаптивна радиация.
Пръстеновидни видове
В биологията пръстеновиден вид е свързана поредица от съседни популации, всяка от които може да се кръстосва със съседните популации. Двата края на веригата се припокриват.
Двете крайни популации в серията са твърде далечно свързани, за да се кръстосват. Такива некръстени, макар и генетично свързани "крайни" популации могат да съществуват едновременно в един и същи регион, като по този начин се затваря "пръстен".
Пръстеновидните видове са важно доказателство за еволюцията: те илюстрират какво се случва с течение на времето, когато популациите се различават генетично. Ричард Докинс отбелязва, че пръстеновидните видове "ни показват само в пространственото измерение нещо, което винаги трябва да се случва във времевото измерение".
Трудно е обаче да се намери прост и ясен пример.
Чайки от вида Larus
Това е класически пример за пръстеновиден вид. Ареалът на тези чайки почти образува пръстен около Северния полюс (който обикновено не се прелита от чайки). Малките черноглави чайки и сребристите чайки са достатъчно различни, за да не се хибридизират обикновено; така че, както се казваше, групата на чайките образува един континуум, освен там, където двете линии се срещат в Европа. Общоприето е обаче, че това не е съвсем вярно, въпреки че подробностите са изключително сложни.
Саламандри Ensatina
Саламандърът Ensatina е пръстеновиден вид в планините около централната калифорнийска долина. Комплексът образува подковообразна форма около планините. Макар че между всяка от 19-те популации около подковата може да има кръстосване, подвидът Ensatina eschscholtzii в западния край на подковата не може да се кръстосва с Ensatina klauberi в източния край. Това е илюстрация на "почти всички етапи в процеса на видообразуване" (Добжански). Ричард Хайтън твърди, че Ensatina е случай на множество видове, а не на континуум от един вид.
Зеленикавата песнопойка
Зеленикавият певец (Phylloscopus trochiloides) има няколко подвида, от които P. t . viridianus е най-познатият в Европа. Това е пръстеновиден вид, чиито популации се разминават на изток и на запад от Тибетското плато, а по-късно се срещат на северната страна. Техните взаимоотношения са доста объркващи.




Симпатрично видообразуване
Симпатричното видообразуване се отнася до формирането на два или повече потомствени вида от един родов вид, които обитават едно и също географско място. Сега се смята, че това е много често срещано явление.
При симпатричното видообразуване видовете се различават, докато обитават едно и също място. Често цитирани примери за симпатрично видообразуване са насекомите, които стават зависими от различни растения-гостоприемници в един и същи район.
Съществуването на симпатричното видообразуване като механизъм за видообразуване беше горещо оспорвано. Хората твърдяха, че доказателствата за симпатично видообразуване са всъщност примери за микрогеографско видообразуване. Като цяло сега това не е предпочитаното обяснение. По-скоро това е резултат от хибридизация между близкородствени видове, последвана от естествен подбор, който въздейства върху потомството на тези кръстоски. Един от широко приетите примери за симпатично видообразуване е този на цихлидите от езерото Набугабо в Източна Африка, за който се смята, че се дължи на полов подбор.
Видообразуване чрез полиплоидизация
Полиплоидията е причина за много бързи събития на видообразуване, тъй като потомството на например тетраплоидни х диплоидни двойки често води до триплоидно стерилно потомство.
Въпреки това не всички полиплоиди са репродуктивно изолирани от родителските си растения и все още може да има генен поток, например чрез триплоидни хибриди х диплоидни двойки, при които се получават тетраплоиди.
Много от съществуващите растителни и повечето животински видове очевидно са претърпели полиплоидизация в еволюционната си история. Размножаването на успешните полиплоидни видове понякога е безполово, чрез партеногенеза. По неизвестни причини много асексуални организми са полиплоидни.
Глог муха
Един пример за еволюция в действие е случаят с мухата глог, Rhagoletis pomonella, която изглежда е подложена на симпатрично видообразуване.
Различните популации на глоговата муха се хранят с различни плодове. Отделна популация се появява в Северна Америка през 19 век, известно време след като са внесени ябълките - неместни видове. Тази популация, хранеща се с ябълки, обикновено се храни само с ябълки, а не с исторически предпочитаните плодове на глога. Настоящата популация, хранеща се с глог, обикновено не се храни с ябълки.
Някои доказателства сочат, че се наблюдава симпатично видообразуване. Глоговите мухи съзряват по-късно през сезона и им отнема повече време да узреят, отколкото на ябълковите мухи; има малко доказателства за кръстосване (изследователите са документирали 4-6% хибридизация).
Появата на новата муха глог е пример за протичаща еволюция.
Хибридизация
В редки случаи се образува нов вид, когато отделни представители на различни видове се чифтосват. Обикновено продуктите от такива хибридни съчетания са безплодни (неплодовити) или относително безплодни и се елиминират от естествения подбор.
Открит е един пример за нов и успешен хибриден вид. Новият вид е хибрид на италианското и испанското врабче, а основните факти са проверени чрез анализ на последователността от ДНК в кръвта им. Птицата живее в Италия, в регион, в който живеят и двата родителски вида. Тя не се размножава с испанското врабче, въпреки че живее с тях рамо до рамо.
Изкуствено видообразуване
Нови видове са създадени чрез отглеждане на домашни животни, но първоначалните дати и методи не са ясни. Например домашните овце са създадени чрез хибридизация и вече не дават жизнеспособно потомство с Ovis orientalis, един от видовете, от които произхождат.
От друга страна, домашните говеда могат да се разглеждат като един и същи вид с няколко разновидности на дивите волове, гаури, якове и т.н., тъй като лесно създават плодовито потомство с тях.
Лабораторни видове
Най-добре документираното създаване на нови видове в лаборатория е извършено в края на 80-те години на миналия век. Уилям Райс и Г. У. Солт развъждат плодови мушици, Drosophila melanogaster, като използват лабиринт с три различни варианта на местообитание, като светло/тъмно и влажно/сухо.
Всяко поколение беше поставено в лабиринта, а групите мухи, които излизаха от два от осемте изхода, бяха отделени, за да се размножават помежду си в съответните групи. След тридесет и пет поколения двете групи и тяхното потомство бяха изолирани репродуктивно поради силните им предпочитания към местообитанията: те се чифтосваха само в рамките на зоните, които предпочитаха, и затова не се чифтосваха с мухи, които предпочитаха другите зони. Историята на подобни опити е описана в Rice and Hostert (1993).
Даян Дод също така успя да покаже как репродуктивната изолация може да се развие от предпочитанията за чифтосване при Drosophila pseudoobscura само след осем поколения, използвайки различни видове храна - нишесте и малтоза.
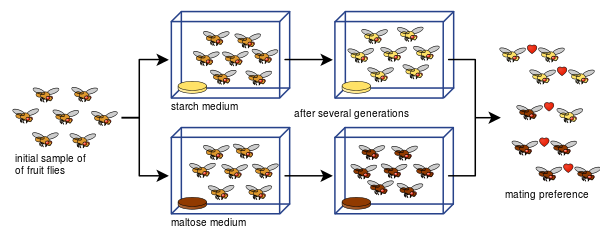
Експериментът на Дод е бил лесен за повторение от много други, включително с други видове плодови мушици и храни.
Подсилване (ефект на Уолъс)
Усилването е процес, при който естественият подбор увеличава репродуктивната изолация.
Тя може да се появи, когато две популации от един и същи вид се разделят и след това отново влязат в контакт. Ако репродуктивната им изолация е била пълна, те вече ще са се превърнали в два отделни несъвместими вида.
Ако тяхната репродуктивна изолация е непълна, по-нататъшното чифтосване между популациите ще доведе до появата на хибриди, които могат да бъдат или да не бъдат плодовити. Ако хибридите са неплодовити или плодовити, но по-малко пригодни от своите предци, тогава няма да има по-нататъшна репродуктивна изолация и видообразуването по същество е настъпило (напр. при конете и магаретата).
Причината за това е, че ако всеки от родителите на хибридното потомство има естествено селектирани черти за своята собствена различна среда, хибридното потомство ще носи черти и от двете и няма да се вписва в никоя от нишите толкова добре, колкото единият от родителите. Ниската пригодност на хибридите би накарала селекцията да предпочете асортативното чифтосване, което би намалило хибридизацията.
Понякога този ефект се нарича ефект на Уолъс по името на еволюционния биолог Алфред Ръсел Уолъс, който в края на XIX век предполага, че той може да бъде важен фактор за видообразуването.
Ако хибридните потомци са по-приспособими от своите предци, популациите ще се слеят отново в един и същ вид в рамките на зоната, в която са в контакт.
Подсилване се изисква за разделяне, когато има "хибридна зона" между две форми на даден вид. Хибридните зони са райони, в които се срещат и кръстосват разминаващи се популации. Хибридното потомство е много често срещано в тези региони, които обикновено се създават от разминаващи се видове, влизащи във вторичен контакт.
Без подсилване двете популации или видове биха се кръстосвали неконтролируемо. Подсилването може да бъде предизвикано при експерименти с изкуствен подбор, както е описано по-горе.
Въпроси и отговори
В: Какво представлява видообразуването?
О: Видообразуването е процесът на формиране на видовете. Той е основна част от еволюционната биология.
В: Какво мисли Дарвин за формирането на видовете?
О: Дарвин е смятал, че повечето видове произлизат директно от вече съществуващи видове, което той нарича анагенеза или "филетична еволюция".
В: Как се гледа на видообразуването като цяло през XX век?
О: През 20-ти век учените смятат, че повечето видове са се образували при разделянето на по-ранни видове, което е известно като кладогенеза. Смятало се е също, че това разделяне е причинено или подпомогнато от изолиращи механизми, като например физическо разделяне.
В: Как последните изследвания промениха разбирането ни за видообразуването?
О: Последните изследвания показаха, че може да се наблюдава хибридизация между сродни видове и чрез тези кръстоски могат да се прехвърлят гени. Това означава, че репродуктивната изолация невинаги е необходима за определянето на даден вид и алопатията (физическото разделяне) може да не е необходима за осъществяването на видообразуване.
Въпрос: Какво общо има физическото разделяне с формирането на нови видове?
О: Смята се, че физическото разделяне между някога съжителстващи видове е основен фактор за формирането на нови видове, което се илюстрира от много примери, разгледани в научната литература.
Свързани статии
Автор
AlegsaOnline.com Видообразуване (специация): как се формират нови видове в еволюцията Leandro Alegsa
URL: https://bg.alegsaonline.com/art/92570
Източници
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833